順遺伝学を用いた連合学習に関わる分子の探索
線虫を用いた研究の醍醐味は、様々な手法が開発されている遺伝学を存分に活用できることです。我々は、順遺伝学を用いて連合学習に関わる分子を同定することに成功しました。線虫の遺伝学については、別項「線虫の分子遺伝学」を参照してください。
−学習異常変異体のスクリーニング−
連合学習は複数の集団を用いてアッセイを行いますので、従来のスクリーニングのように1匹ずつ表現型を同定することはできません。そこで図1のようにF2を複数プレートに分けて、それぞれNaCl+条件付け後にNaClへの化学走性アッセイを行いました(学習アッセイ)。30分後に、NaClへ誘引された個体を回収しました。多くの野生型線虫は、学習によりNaClを忌避するようになりますので、この集団の中における変異体の存在比は、アッセイ前と比較して高くなることが期待されます。回収してきた虫をNGMで生育し、それらの次世代についても同様のアッセイを行いました。このような選択を繰り返し行うことで、全個体中の変異体が占める割合(存在比)は高くなると予想されます。実際にスクリーニングと同時並行する形で、ins-1学習変異体を用いて疑似的なスクリーニングを行ったところ、回数を重ねるにつれてins-1の存在比が飛躍的に高くなりました(表1)。このようなスクリーニングにより、最終的に5系統(JN401〜JN405)の変異体候補株を獲得しました(図2)。
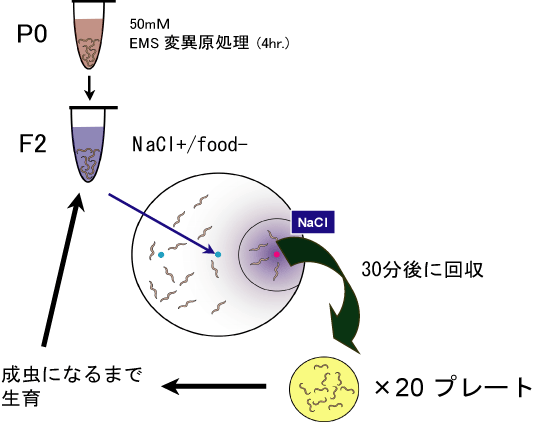
図1 スクリーニングの概略 |
表1 ins-1 変異体の濃縮
(疑似スクリーニング)
|
Wild type |
ins-1 |
Ratio |
| Before assay |
1200 |
23 |
2% |
| After 1st. assay |
27 |
10 |
27% |
| After 2nd. assay |
5 |
116 |
96% |
| After 3rd. assay |
1 |
>50 |
>98% |
|
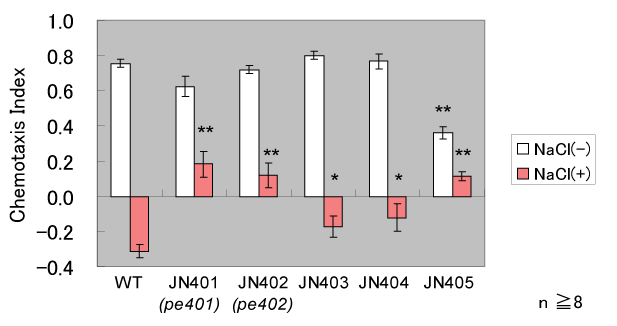 |
図2 獲得した学習異常変異体
我々の単離した変異体(JN401〜405)は、野生株(WT)と比べてNaClと飢餓による条件付け後の 化学走性指数(NaCl(+), 赤い棒)が高い値を示す。すなわち、条件付け後の化学走性の低下が
異常であるといえる。縦軸は化学走性指数(Chemotaxis Index)={(塩に 寄ってきた線虫の個体数)-(塩から逃げた線虫の個体数)}/(全線虫の個体数)}
をあらわす。 |
−表現型の詳細な解析−
これらの変異体の通常状態における塩や餌に対する応答について調べました。特にpe401とpe402という2つの変異体では、塩や餌に対する反応が、ほぼ正常であることが明らかになりました。つまり、NaClに対する感受性が低下したということではなく、条件付けを行った後の可塑的な変化に対してのみ異常を示す、学習異常変異体であると考えられます。
−原因遺伝子の同定−
遺伝学的マッピング解析により、それぞれの原因遺伝子の同定を試みました。その結果、pe402変異体において、scd-2という受容体型チロシンキナーゼをコードする遺伝子領域に1.3kbほどの欠失があることを発見しました(図3)。連合学習のプロセスでSCD-2がどのように機能するのかについては、現在九州大学の研究室と共同研究を進めています。ちなみに、我々はINS-1(インシュリン様分子)の受容体と推定されているDAF-2の変異体も、ins-1変異体と同様、学習に欠損を示すことを明らかにしていますが(詳しくはこちらを参照)、このDAF-2もやはり受容体型チロシンキナーゼです。
また最近、pe401変異体の原因遺伝子として、新奇の細胞接着関連因子を同定しました。両者ともに神経系における機能など不明な点が多いのですが、多細胞生物で広く保存されている因子であり、多くの生物種で重要な働きを示すものと期待されます。
(池田 大祐)
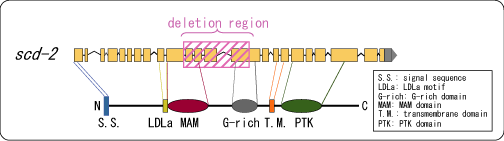
図3 scd-2遺伝子構造と同定された欠失(deletion)領域
|