連合学習における下流因子の検索
近年では神経可塑性または行動可塑性を制御する様々な分子メカニズムが明らかになってきました。例えば、プレシナプスにおける神経伝達物質分泌の制御やポストシナプスにおける神経伝達物質受容体の制御などを介したシナプス強度の可塑的変化のメカニズムは広く知られている現象です。
当研究室では、インスリンシグナル伝達経路が行動可塑性を制御することを見出しています(詳しくはこちらを御参照ください)。いったいこのシグナル伝達経路が神経細胞レベルでどの様な制御をしているのでしょうか?現在までのところ、INS-1(インスリン)により活性化されたDAF-2(インスリン受容体)はAGE-1(PI3キナーゼ)を活性化し、その下流でPDK-1やAKT-1などのタンパク質リン酸化酵素が活性化することが示唆されています。我々は、NaClと飢餓による連合学習においてインスリン/PI3キナーゼシグナル伝達経路の下流で働く分子を同定するために以下に示すような方法によりスクリーニングを行っています。
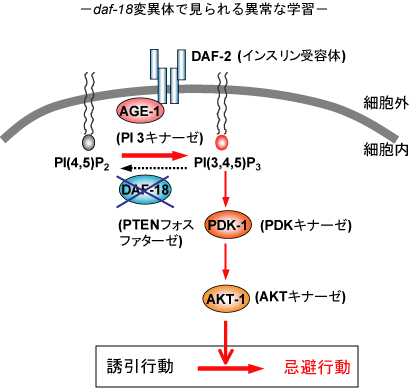 |
| daf-18変異体は飢餓とNaClによる条件付けをしなくてもNaClに対して忌避応答を示す。この変異体では、DAF-18の機能が低下しているためにDAF-2が活性化されなくても、AGE-1の触媒産物であるであるPI(3,4,5)P3のレベルが上昇している。このことから、daf-18変異体では学習後の野生株と同様な状態であり、条件付けをする前から「異常な学習」をしていると考えられる。 |
上の図に示しますようにage-1 (PI3キナーゼ)の逆反応を触媒する酵素であるdaf-18 (PTENホスファターゼ)の変異体では、通常の状態において(飢餓とNaClによる条件付けをする前から)NaClに対して忌避応答を示します。このことは、daf-18の機能低下によりage-1の相対的な活性が上昇しているために、通常状態において「異常な学習」をしていると解釈することができます。
そこで、daf-18変異体の「異常な学習」を抑圧(サプレス)する変異をサプレッサースクリーニングにより探索しています。このような変異体ではage-1の下流で連合学習を制御している分子の機能が低下していると期待できます。(例えば、daf-18変異とage-1の下流で働くことがわかっているakt-1の変異との2重変異体は通常状態において野生株と同様のNaClへの誘引反応を示します。これは、akt-1変異がdaf-18変異体の表現型を抑圧(サプレス)したといえます。
具体的なスクリーニング手順としては、まずdaf-18変異体に対して変異原処理を行いました。その後、(条件付けを行わない通常の)化学走性アッセイを行い、学習異常変異体スクリーニングの時と同じ要領で、NaClへ誘引された個体を回収しました。このような濃縮を行った結果、daf-18の異常な学習を抑圧した株を23系統、そのうち学習欠損を示す株を13系統獲得しました。(遺伝学的スクリーニングの一般的な原理を知りたい方はこちらを参照してください。)
今後、これら変異体の原因変異を同定し、インスリン/PI3キナーゼシグナル伝達経路の下流で働く分子を明らかにしていく予定です。
(安達健・富岡征大)
|